crispr-cas系统是原核生物抵御外来噬菌体入侵的一种获得性免疫防御系统。根据cas效应蛋白的组成,crispr-cas系统分为class 1和class 2。class 1系统(进一步分为i、iii和iv型)的效应蛋白由多个亚基的效应复合物组成,而class 2系统(ii、v和vi型)的效应蛋白为单个蛋白。在这些类型中,iii型crispr-cas系统因其复杂精密的免疫机制和对入侵者rna以及dna的切割而备受关注。iii型系统识别入侵的rna分子,并进一步分为iii-a到iii-f型。
makarova等在2019年通过生物信息学分析报道了iii-e型crispr-cas系统1,在该系统中,4个cas7结构域和一个类似csm2/cmr5的小亚基(cas11)融合成一个大的蛋白质,被命名为gramp。在iii-e型crispr-cas系统的基因cluster中,iii型系统标志性的cas10以及csm4/cmr5基因缺失,表明它的免疫机制会有所不同。后续研究发现,iii-e型系统能将前体crrna加工成成熟的crrna,并将靶rna以6个核苷酸为间隔进行切割,且没有rna旁切效应,有望成为一种高特异性的rna编辑工具2, 3。而且有趣的是,iii-e型的效应蛋白gramp能稳定地与caspase样蛋白酶tpr-chat形成一个更大的复合物(被命名为craspase),它被推测是可通过iii-e型效应蛋白识别靶rna而被激活的蛋白酶。然而,该蛋白酶活性没有被证实,tpr-chat的底物也是未知的。此外,iii-e型crispr-cas系统的免疫机制也仍然是个谜。
2022年10月27日,北京化工大学冯越教授课题组与清华大学杨茂君教授课题组合作在molecular cell在线发表了题为target rna activates the protease activity of craspase to confer antiviral defense的研究论文。该项工作报道了来自candidatus "scalindua brodae"的gramp-crrna及craspase的一系列结构,发现ctr (cognate target rna)诱导craspase中的tpr-chat发生显著的构象变化,并揭示了ctr激活craspase的蛋白酶活性切割底物蛋白csx30,从而触发细菌的顿挫感染(abortive infection)作为iii-e型系统的抗病毒策略。

课题组首先通过对gramp-crrna以及gramp-crrna-tr (target rna) 结构的分析,发现gramp中d547以及d698/d806分别负责靶rna中site 1和site 2的切割。值得注意的是,gramp的cas7.4结构域中的插入结构域(insertion domain)在电镜数据中的密度较差,并且insertion domain缺失的gramp突变体显示出对靶rna切割活性的缺失,以及较弱的pre-crrna加工活性。这表明插入结构域可能参与了gramp的前体crrna加工,这一点与来自desulfonema ishimotonii物种的gramp存在差异4,这可能反映了不同物种iii-e型效应蛋白的机制差异。
那么,tpr-chat的蛋白酶活性是否能被激活以及蛋白酶靶向的底物是什么?课题组接下来把研究的重点放在了craspase上,对craspase自身以及与tr/ctr/ntr (non-cognate target rna)结合后四种状态下的结构进行了解析。结构比对发现,craspase与ctr结合(即靶向“非我”rna)后会引起tpr-chat中包括活性位点在内的区域显著的构象变化,而tr/ntr则不能引起这样的变化。这表明,craspase中tpr-chat的蛋白酶活性可能被ctr的结合所激活。
为了寻找tpr-chat的底物,课题组对iii-e型的基因cluster进行了分析,在该cluster中通常有三个保守的辅助基因,分别是csx30、csx31和一个sigma因子e(σe)编码sbrpoe。蛋白酶活性实验表明,在ctr存在的情况下,craspase可以切割csx30,但不能切割sbrpoe,并且craspase对csx30的蛋白酶活性不能在tr或ntr存在的情况下被激活。由于csx31单独不能纯化到均一状态,所以没有对单独的csx31蛋白进行酶切检测。同时,他们还发现csx30、csx31和sbrpoe可形成一个复合物,激活的craspase也可以该复合物作为底物,切割其中的csx30蛋白。
为了研究ctr激活的csx30切割的生物学意义,课题组将编码gramp、tpr-chat、csx30、csx31和sbrpoe的基因以及含有针对dsdna噬菌体lambda(λ)基因转录物序列的合成crispr array转化到大肠杆菌中,进行λ噬菌体侵染实验以及细菌体内诱导生长实验。实验结果表明,该系统能够赋予大肠杆菌抗λ噬菌体的活性,并且采用的机制为顿挫感染。同时,craspase识别ctr而触发的对csx30的切割对细胞有毒性,切开的csx30的两段与csx31和sbrpoe在大肠杆菌中共表达时会抑制大肠杆菌的生长。
基于以上的研究,课题组提出了iii-e型免疫系统的分子机制模型,即craspase中的tpr-chat本身处于失活状态,而噬菌体来源的核酸转录出的rna结合在craspase后诱导tpr-chat的构象变化并激活其活性,tpr-chat识别底物csx30并进行切割,从而在csx31和sbrpoe存在下引发细菌的顿挫感染(图1)。然而,csx30的切割在csx31和sbrpoe存在的情况下共同诱发细胞死亡的详细机制仍有待后续研究。
北京化工大学博士研究生刘茜、清华大学博士后张来幸、北京化工大学博士研究生王浩、硕士研究生修瑀,黄玲,高政宇为本论文的共同第一作者。冯越教授为本文的通讯作者和 lead contact,清华大学杨茂君教授和张来幸博士为本文的共同通讯作者,北京化工大学为第一完成单位。
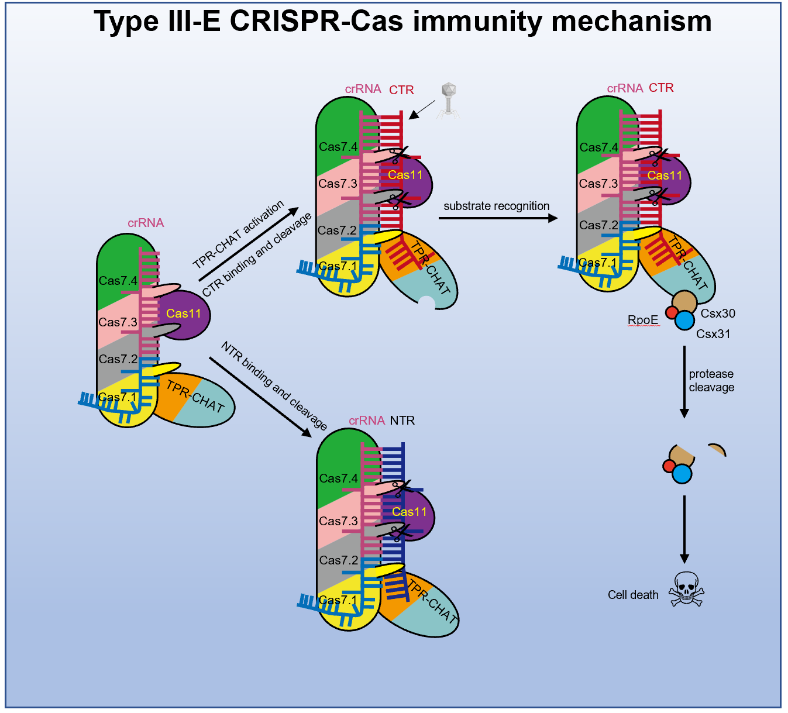
图1. iii-e型crispr-cas系统的免疫机制模型
参考文献
1.makarova, k.s. et al. evolutionary classification of crispr-cas systems: a burst of class 2 and derived variants. nat rev microbiol18, 67-83 (2020).
2.ozcan, a. et al. programmable rna targeting with the single-protein crispr effector cas7-11. nature597, 720-725 (2021).
3.van beljouw, s.p.b. et al. the gramp crispr-cas effector is an rna endonuclease complexed with a caspase-like peptidase. science373, 1349-1353 (2021).
4.kato, k. et al. structure and engineering of the type iii-e crispr-cas7-11 effector complex. cell185, 2324-2337 e2316 (2022).